开花时间调节剂FCA和FPA在RNA介导的染色质沉默中的广泛作用
Isabel Bauml; urle,1* Lisa Smith,2dagger; David C. Baulcombe,2Dagger; Caroline Dean1*
摘要:RRM结构域蛋白FCA和FPA以前被认为是拟南芥开花时间的调节因子。我们发现它们是RNA介导的基因组中一系列位点染色质沉默所必需的。在某些靶位点,FCA和FPA促进DNA不对称甲基化,而在另一些靶位点,FCA和FPA的作用与DNA甲基化平行。雌配子体发育和早期胚胎发育特别容易发生FCA和FPA的功能紊乱。我们认为FCA和FPA调节单拷贝和低拷贝基因的染色质沉默,并以基因座依赖的方式与典型的小干扰RNA导向的DNA甲基化途径相互作用以调节共同的靶点。
许多生物体中的H-染色质的特征是广泛的DNA甲基化和组蛋白修饰(1)。植物在CG、CNG(N=任意核苷酸)和CHH(H=A、C或T)序列上下文中显示胞嘧啶甲基化。在拟南芥中,小干扰RNA(SiRNAs)参与定位和维持这些染色质修饰的过程,需要RNADEPENDENT RNA POLYMERASE2(RDR2)、DICER-LIKE3(DCL3)、ARGONAUTE4(AGO4)和两个RNA聚合酶IV亚型,Pol IVa和b(2-9)。
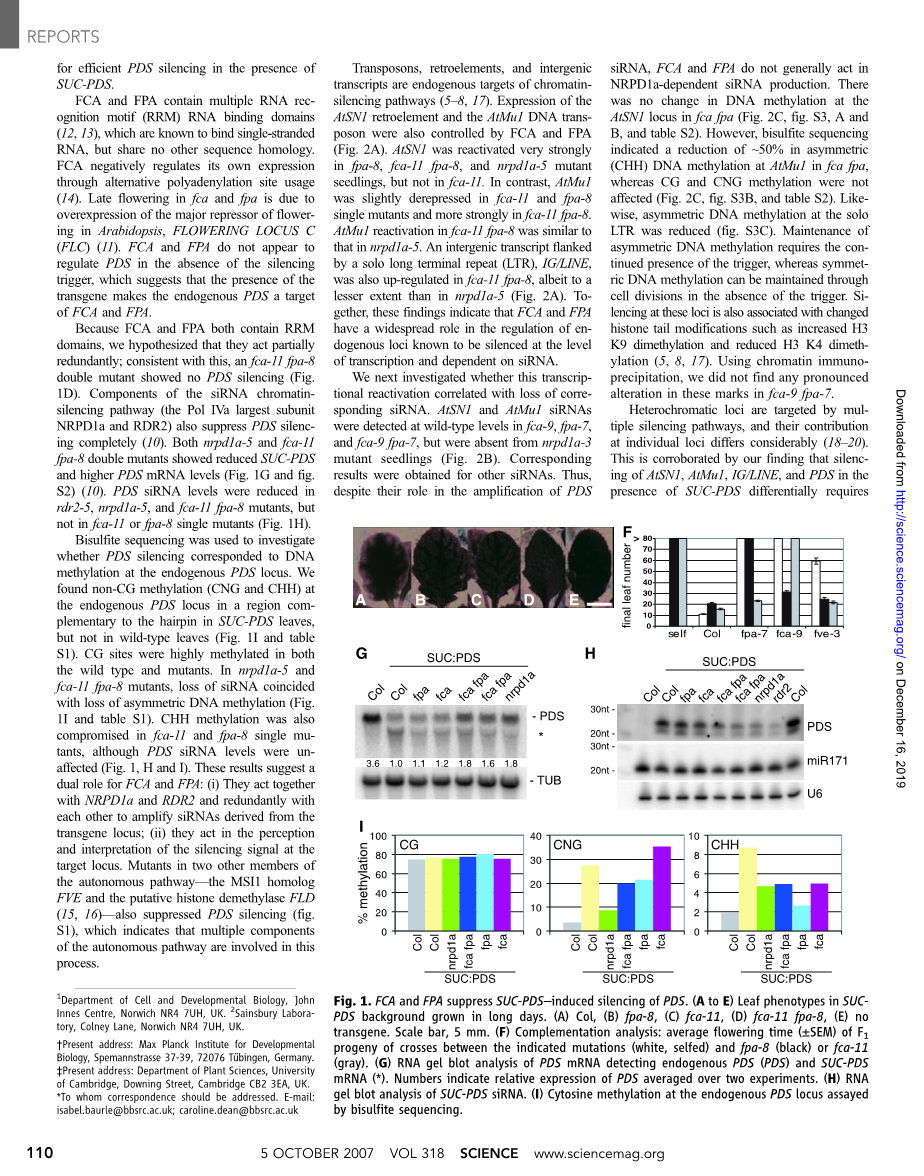
为了确定siRNA介导的染色质沉默所需的其他组件,我们使用了一种报道基因系统,其中拟南芥八氢番茄红素去饱和酶(PDS)基因响应同源反向重复(SUCPDS)而被沉默(10)。两个部分抑制PDS沉默的突变体(图1,A,B,C和E)显示了后期开花,可通过春化来逆转。沉默和开花表型共分离,并且突变定位于染色体2和4。开花表型表明,FPA和FCA是自主途径的两个成员(11),涉及到那些基因组区域。测序显示FPA(Trp98*,G至A,fpa-8)和FCA(Gln537*,C至T,fca-11)中的过早终止密码子。通过与先前已知的开花突变体(fca-9,fpa-7和fve-3;图1F)进行互补分析确认了开花缺陷,该突变体也显示PDS沉默(图S1)。因此,需要FCA和FPA在存在SUC-PDS的情况下实现有效的PDS沉默。
FCA和FPA包含多个RNA识别基序(RRM)RNA结合结构域(12、13),其中RNA不共享其他序列同源性。FCA通过替代性聚腺苷酸化位点的使用来负面调节其自身的表达(14)。fca和fpa的晚期开花是由于拟南芥开花的主要阻遏物FLOWERING LOCUS C(FLC)的过度表达所致(11)。在没有沉默触发的情况下,FCA和FPA似乎没有调节PDS,这表明转基因的存在使内源性PDS成为FCA和FPA的特征。
由于FCA和FPA都包含RRM域,因此我们假设它们的作用是部分冗余的。与此相一致,一个fca-11 fpa-8双重突变体显示没有PDS沉默(图1D)。 siRNA染色质沉默途径的组成部分(Pol IVa是最大的亚基NRPD1a和RDR2)也完全抑制了PDS沉默(10)。nrpd1a-5和fca-11 fpa-8双重突变体均显示出降低的SUC-PDS和较高的PDS mRNA水平(图1G和图S2)(10)。PDS siRNA水平在rdr2-5,nrpd1a-5和 fca-11 fpa-8突变体中降低,但在fca-11或fpa-8单突变体中未降低(图1H)。
亚硫酸氢盐测序用于研究PDS沉默是否与内源PDS基因座处的DNA甲基化相对应。我们在SUC-PDS叶片中与发夹互补的区域的内源PDS基因座中发现了非CG甲基化(CNG和CHH),而在野生型叶片中则没有(图1I和表S1)。在野生型和突变体中,CG位点都高度甲基化。在nrpd1a-5和fca-11 fpa-8突变体中,siRNA的丢失与不对称DNA甲基化的丢失同时发生(图1I和表S1)。尽管PDS siRNA水平不受影响,但在fca-11和fpa-8单个突变体中CHH甲基化也受到损害(图1,H和I)。这些结果表明了FCA和FPA的双重作用:(i) 与NRPD1a和RDR2相互冗余,相互冗余地扩增源自转基因基因座的siRNA。(ii)它们在目标基因座的沉默信号的感知和解释中起作用。自主途径的其他两个成员(MSI1同系物FVE和推定的组蛋白脱甲基酶FLD(15、16)的突变体也抑制了PDS沉默(图S1),这表明该过程涉及自主途径的多个组成部分。
转座子,逆转录元件和基因间转录本是染色质沉默途径的内源性靶标(5-8、17)。FCA和FPA也控制了AtSN1元件和AtMu1 DNA转座子的表达(图2A)。在fpa-8,fca-11,fpa-8和nrpd1a-5突变苗中,AtSN1的激活非常强烈,但在fca-11中却没有。相反,AtMu1在fca-11和fpa-8单个突变体中略微降低,而在fca-11 fpa-8中则更弱。 fca-11 fpa-8中的AtMu1重新激活与nrpd1a-5中的类似。在fca-11 fpa-8中,侧翼是一个长末端重复序列(LTR)的基因间转录物也被上调,其程度要小于nrpd1a-5(图2A)。迄今为止,这些发现表明FCA和FPA在调节内源基因座中具有广泛的作用,该内源基因座已知在转录水平上是沉默的并且依赖于siRNA。
接下来,我们研究了这种转录重新激活是否与相应的siRNA丢失相关。在fca-9,fpa-7和fca-9 fpa-7中以野生型水平检测到AtSN1和AtMu1 siRNAs,但在nrpd1a-3突变苗中却没有(图2B)。其他siRNA也获得了相应的结果。因此,尽管它们在PDS siRNA的扩增中发挥作用,但FCA和FPA通常不依赖于NRPD1a的siRNA产生。 fca fpa中AtSN1位点的DNA甲基化没有变化(图2C,图S3,A和B和表S2)。但是,亚硫酸氢盐测序表明fca fpa中AtMu1处的不对称(CHH)DNA甲基化减少了约50%,而CG和CNG甲基化未受影响(图2C,图S3B和表S2)。同样,单独LTR处的不对称DNA甲基化也降低了(图S3C)。维持不对称DNA甲基化需要触发器的持续存在,而对称DNA甲基化则可以在不存在触发器的情况下通过细胞分裂来维持。这些位点的沉默也与组蛋白尾部修饰的改变有关,例如增加的H3 K9二甲基化和减少的H3 K4二甲基化(5、8、17)。使用染色质免疫沉淀,我们在fca-9 fpa-7中没有发现这些标记的任何明显改变。
图1. FCA和FPA抑制SUC-PDS引起的PDS沉默。(A到E)SUCPDS背景中的叶片表型生长很长时间。 (A)Col,(B)fpa-8,(C)fca-11,(D)fca-11 fpa-8,(E)无转基因。比例尺,5毫米。(F)互补分析:所示突变(白色,自交)和fpa-8(黑色)或fca-11(灰色)之间杂交的F1子代的平均开花时间(plusmn;SEM)。 (G)检测内源性PDS(PDS)和SUC-PDS mRNA(*)的PDS mRNA的RNA凝胶印迹分析。数字表示在两个实验中平均的PDS相对表达。(H)SUC-PDS siRNA的RNA凝胶印迹分析。(I)通过亚硫酸氢盐测序测定的内源PDS基因座处的胞嘧啶甲基化。
异色基因座是由多个沉默途径靶向的,它们对单个基因座的贡献差异很大(18-20)。我们的发现证实了这一点,即在存在SUC-PDS的情况下,对AtSN1,AtMu1,IG/LINE和PDS的沉默要求不同FCA和FPA。PDS沉默通过依赖于siRNA染色质沉默途径和fca fpa的机制与靶DNA甲基化和siRNA产生相关。在fca fpa突变体中,AtMu1和IG/LINE的抑制与DNA甲基化的丧失相符,但与siRNA的丧失相符,而在siRNA染色质沉默途径的突变体中都缺失。尽管在fca fpa中AtSN1的激活作用强得多,但DNA甲基化和siRNA积累均不受影响。我们的发现与这样的想法是一致的,即转录可以在DNA甲基化的情况下重新激活,这是针对吗啡分子1(mom1)突变的证据(19,21)。尽管有这种相似性,FCA和FPA似乎不太可能与MOM1一起起作用,因为在mom1中AtSN1和AtMu1并未受到错误调节(22)。
为了研究FCA和FPA如何与染色质siRNA扩增途径(包括Pol IVa,RDR2和DCL3)相关,我们分析了双突变体中沉默的释放(图2D)。所有双突变体均显示出比任何单个突变体高得多的AtSN1和AtMu1激活,这表明FCA和FPA并不在siRNA扩增途径的下游起作用,而是并行发挥作用。同样,相对于单个突变体(图S3D)中的任何一个,在fve nrpd1a双重突变体中转座子的激活都大大增强了。令人惊讶的是,尽管FCA对于野生型的AtSN1沉默是必不可少的,但nrpd1a,rdr2或dcl3突变体背景下FCA的缺失大大增强了AtSN1沉默的释放。
我们的发现预测,fca fpa突变体中DNA甲基化的扰动将以不同方式影响目标基因座的重新激活。在AtSN1,FCA和FPA的作用与DNA甲基化脱钩,在存在DNA甲基化抑制剂5-氮杂-脱氧胞苷(aza-dC)的情况下,沉默的损失会增加。相反,在AtMu1,fca fpa突变体显示出DNA甲基化降低,该抑制剂的附加作用很小。我们的结果(图3A和表S3)与该预测一致,因为就AtSN1激活而言,fca-9 fpa-7突变体对azadC的敏感性比野生型高,而对AtMu1的敏感性比野生型低。重新激活。同样,当暴露于aza-dC时,野生型或fca-9幼苗的发育并非异常且fpa-7幼苗的发育仅非常轻微的异常时,fca-9 fpa-7幼苗的发育受到强烈干扰。(图3B和表S4)(23)。
fca fpa双重突变植物开花较晚,但在其他情况下则基本正常。但是,仔细检查fca-11 fpa-8角果后发现,约20%的正在发育的种子已中止,约70%的胚珠未开始发育(图S4A和表1)。当用野生型花粉对双重突变体进行授粉时,没有种子流产,但仍有大量未发育的种子持续存在。这一发现表明,胚胎致死性是合子的,而未发育的种子表型是由母体的基因型引起的。当用野生型花粉对fca/fca FPA/fpa胚珠进行授粉时,34%的种子似乎未发育(表1)。显微镜检查成熟胚珠未发现任何异常(图S4,B和C),这表明雌配子体的基因型决定了未发育的种子表型。
剩余内容已隐藏,支付完成后下载完整资料

英语原文共 5 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[275536],资料为PDF文档或Word文档,PDF文档可免费转换为Word
课题毕业论文、外文翻译、任务书、文献综述、开题报告、程序设计、图纸设计等资料可联系客服协助查找。

