基因编辑减数分裂和受精基因获得杂交水稻无性种子
作者:Chun Wang1, Qing Liu1, Yi Shen2, Yufeng Hua1, Junjie Wang1, Jianrong Lin1, Mingguo Wu1, Tingting Sun1, Zhukuan Cheng2 , Raphael Mercier 3 and Kejian Wang 1
单位:1:State Key Laboratory of Rice Biology, China National Rice Research Institute, Chinese Academy of Agricultural Sciences, Hangzhou, China.
2 :State Key Laboratory of Plant Genomics, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China.
3 :Institut Jean-Pierre Bourgin, INRA, AgroParisTech, CNRS, Universiteacute; Paris-Saclay, Versailles, France.
专业:生物技术 学生姓名:罗灶霞
指导老师姓名:黄小平
摘要:育种家利用杂种优势培育优良的高产作物品系,但由于遗传分离,在后代中丧失了有益的表型。通过种子进行无性系繁殖可以使F1杂种自繁。本文报告了一种通过种子实现F1水稻杂交种无性繁殖的策略。我们通过对REC8、PAIR1和OSD1减数分裂基因的CRISPR-Cas9多重基因组编辑,固定F1杂交稻杂合度,获得无性系二倍体配子和四倍体种子。基因(参与受精)可诱导杂交水稻单倍体种子的形成。最后,我们通过对杂交稻中的4个基因(REC8、PAIR1、OSD1和MTL)的同时编辑,将杂合性固定和单倍体诱导结合起来,获得了能够通过种子进行无性繁殖的植株。该方法的应用可以使广泛的优良F1杂交作物自繁殖。
遗传距离较远的个体杂交后代由于杂种优势,相对于纯合亲本,活力增加。杂种优势已被广泛应用于农业,以提高作物的生产力和适应性[1,2]。然而,对许多作物来说,杂交种子的生产是非常昂贵的。合成无融合生殖已被认为是固定F1杂交种作物品种杂种优势的途径[3]。无融合生殖是一种无性繁殖策略,其后代是通过种子产生而不经过减数分裂或受精。虽然无融合生殖在许多开花植物类群中已有描述[4],但在作物中尚未见报道。在拟南芥和水稻中,3个介导重要减数分裂过程的基因的联合突变创造了一个名为MiMe(有丝分裂而非减数分裂)的基因型,在该基因型中减数分裂被有丝分裂样分裂所取代,从而产生雌雄克隆二倍体配子[5,6]。然而,MiMe植株的自交受精使每代倍性提高一倍。将拟南芥MiMe与CENH3介导的染色体消除系杂交产生克隆二倍体后代[7]。然而,这个系统仍然依赖于杂交不同的植物,CENH3介导的染色体消除似乎不太可能转移到其他物种上[8]。因此,我们着手设计一种在自交F1杂种中广泛适用的杂种优势固定方法。
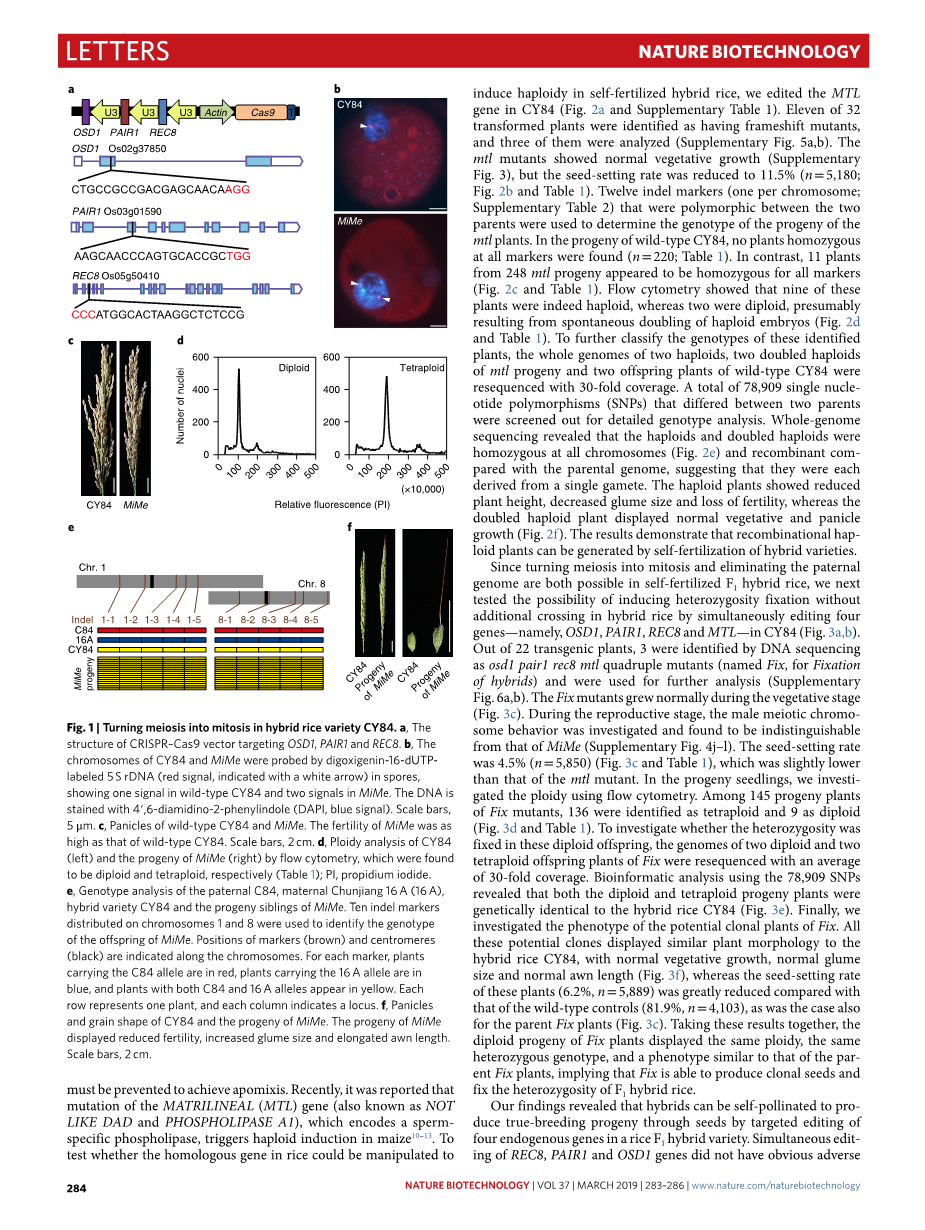
首先,为了验证MiMe技术在杂交稻品种上的可行性,我们从粳稻雄性不育系春江16A(16A)和籼粳中间型杂交稻父本C84杂交后代中选出了优良的亚种间杂交稻lsquo;春优84rsquo;(CY84)(附图1)。为了保证MiMe在CY84杂交背景下的快速生成,我们利用我们前期开发的多重CRISPR - Cas9系统[9]同时编辑了REC8、PAIR1和OSD1基因(图1a和附表1)。32株初代转化植株中有7株被鉴定为移码三突变体,其中3株被详细分析(附图2a,b)。三重突变体(MiMe)在生长情况和形态特征上都无法与野生型CY84区分(附图3)。为了检验减数分裂是否已转变为有丝分裂样分裂,我们研究了野生型和MiMe植物的雄性减数分裂染色体行为。在野生型CY84(补图4a-f)中,12对二价体在终变期分散,在中期Ⅰ沿赤道板排列,12对同源染色体在后期Ⅰ分离,第二次减数分裂后产生四分体。在MiMe(附图4g-i)中,在终变期时发现24个单价体,并在中期Ⅰ时对齐。在后期Ⅰ中,24对染色单体分成两组,并产生二分体,表明减数分裂变成有丝分裂。然后利用5S rDNA特异性探针对水稻11号染色体进行荧光原位杂交分析,检测了MiMe孢子的倍性。在CY84孢子(n=30)中只观察到一个信号,而在MiMe孢子(n=40,图1b)中则观察到两个信号,表明MiMe中产生了二倍体配子。我们还对MiMe突变体的育性进行了研究,发现MiMe中结实率为81.2 % (n=4,043),与野生型(79.1%,n=3,876)相当(图1c和表1),说明这三个基因同时编辑对该杂交种的育性没有明显影响。利用流式细胞仪对MiMe植株后代的倍性进行了研究,发现所有植株(n=123)均为四倍体植株(图1d和表1)。此外,这些后代植株(n=123)完全保留了CY84亲本对10个被测插入缺失标记(图1e和附表2)的杂合性。与野生型相比,MiMe自交后代表现出育性降低、籽粒增大、芒长拉长等特点,均为四倍体水稻的特征(图1f)。这些结果表明,利用CRISPR - Cas9基因组编辑技术可以将MiMe表型快速导入杂交水稻品种中。
附图1. 亚种间杂交水稻品种春优84(CY84)及其亲本的形态。
母本为晚粳不育系lsquo;春江16Arsquo;(简称16A ),父本为广亲和的籼粳中间型恢复系lsquo;C84rsquo;。用保持系春江16B ( 16B )与近等基因16A杂交,获得雄性不育16A种子。比例尺,5thinsp;cm。
图1. 杂交水稻品种CY84减数分裂转化有丝分裂图
(a). 靶向OSD1、PAIR1和REC8的CRISPR - Cas9载体的结构 (b). 利用地高辛素-16 - dUTP标记孢子中5thinsp;S rDNA ( 红色信号 ,用白色箭头表示)对CY84和MiMe染色体进行探测,在野生型CY84中显示1个信号,在MiMe中显示两个信号。DNA用4′,6 -二脒基-2 -苯基吲哚(DAPI ,蓝色信号 )染色。比例尺,5 mu; m (c).野生型CY84和MiMe的角质层。MiMe的育性与野生型CY84一样高。比例尺,2thinsp; cm (d).利用流式细胞仪对CY84 (左)和MiMe (右)子代进行倍性分析,分别发现为二倍体和四倍体(表1 );PI,碘化丙啶 (e).父本C84、母本春江16A ( 16thinsp;A )、杂交品种CY84和MiMe后代兄弟姐妹的基因型分析。利用分布在第1条和第8条染色体上的10个插入缺失标记鉴定了MiMe后代的基因型。标记(棕色)和着丝粒(黑色)的位置沿染色体指示。对于每个标记,携带C84等位基因的植株呈红色,携带16thinsp;A等位基因的植株呈蓝色,同时携带C84和16thinsp;A等位基因的植株均呈黄色。每行代表一个植物,每列代表一个基因座 (f).CY84和MiMe后代的穗粒形状。MiMe的后代表现为育性降低,颖片大小增加,芒长拉长。比例尺,2thinsp;cm。
附图2. MiMe的基因组编辑结果。
( a ). MiMe的T0转基因植株的基因型。字母A - C分别代表OSD1、PAIR1和REC8基因。上例字母表示没有突变,而下例字母表示移码突变;较低的case*表示非移码突变。( b ) . MiMe- # 7、MiMe- # 8和MiMe- # 21的OSD1、PAIR1和REC8靶点周围突变的测序分析。野生型(WT)的前间区序列相邻基序和靶序列分别用红色和蓝色表示。符号-表示已删除的核苷酸。
附图3. 野生型CY84、MiMe和MTL的形态。
MiMe和MTL突变体表现正常营养生长。比例尺,5thinsp;cm。
附图4. 野生型CY84、MiMe和Fix花粉母细胞中雄性减数分裂的染色体扩展。
( a-f ) CY84 ( n = 32 ).(a)具有12个对齐的二价体的中期Ⅰ. ( b )后期Ⅰ. ( c ) 核分裂末期I. (d )中期Ⅱ. ( e )后期Ⅱ. ( f ) 核分裂末期Ⅱ. ( g-i ) MiMe ( n = 45 ). ( g )具有24个对齐单价的中期I。( h )具有24对染色单体的后期I.(i)核分裂末期 I . ( j-l )Fix( n = 52 )。( j ). 具有24个对齐单价的中期I.(k)具有24对染色单体的后期I.(l)核分裂末期I .比例尺,5 m。
图2. 通过编辑杂交水稻品种CY84中的MTL基因产生单倍体诱导系。
(a). 靶向MTL的CRISPR-Cas9载体 . 的结构示意图. (b). CY84背景下野生型和MTL的圆锥花序。MTL的育性下降;白色箭头表示流产的种子,红色箭头表示可育的种子。比例尺,2thinsp;cm. (c). 裁剪后的凝胶用12个插入缺失标记(每条染色体1个)表示单倍体、双倍体( DH )和重组自交系二倍体( RID )的基因型。MTL后代所有标记纯合的植株鉴定为单倍体或双倍体. (d). 流式细胞仪对单倍体和DH进行倍性分析(表1 );PI,碘化丙啶. (e). 单倍体、DH和RID植株全基因组测序。12块代表12条染色体。C84等位基因的SNP为红色,16thinsp;A等位基因的SNP为蓝色,两种等位基因共存于黄色. (f). 野生型CY84和MTL后代的圆锥花序,包括RID、单倍体和DH植物。比例尺,2cm。
MiMe克隆配子参与正常的自交受精,产生双倍性的后代。必须防止这种倍性倍增,以达到无融合生殖。近来有报道称,编码精子特异性磷脂酶的MATRILINEAL(MTL)基因(又称NOT LIKE DAD和磷脂酶A1)突变引起玉米单倍体诱导[10 - 13]。为了验证水稻同源基因是否可以在自交杂交水稻中操纵诱导单倍体,我们编辑了CY84中的MTL基因(图2a和附表1)。32株转化植株中有11株被鉴定为移码突变体,并对其中3株进行了分析(附图5a,b)。MTL突变体植株营养生长正常(附图3),但结实率降低到11.5%(n=5,180;图2b和表1)。利用12个在两个亲本间具有多态性的内模标记(每条染色体1条;附表2)来确定MTL植株后代的基因型。在野生型CY84的后代中,所有标记均未发现纯合植株(n=220;表1)。相比之下,来自248MTL子代的11株植物对所有标记似乎都是纯合的(图2c和表1)。流式细胞术显示,这些植物中9株确实是单倍体,2株是二倍体,推测可能是单倍体胚胎自发加倍所致(图2d和表1)。为了进一步对这些鉴定植株的基因型进行分类,我们对两个单倍体、两个MTL子代双倍体和两个野生型CY84子代植株的全基因组进行了30倍复盖率的测序,共筛选出78909个双亲本间差异的单核苷酸多态性位点,对其进行详细的基因型分析。全基因组测序显示,单倍体和双倍单倍体在所有染色体上都是纯合的(图2e),与亲本基因组相比都是重组的,表明它们各自来源于一个配子体。单倍体植株表现为株高降低,颖片变小,育性丧失,而双倍单倍体植株表现为正常的营养生长和穗生长(图2f)。结果表明,杂交品种自交可产生重组单倍体植株。
附图5. MTL的基因组编辑结果
( a )转MTL基因T0植株的基因型。D表示无突变,d表示移码突变,d *表示MTL的非移码突变。( b ) mtl- # 1、mtl- # 2和mtl- # 3 MTL靶点周围突变的测序分析。野生型( WT )的前间区序列相邻基序和靶序列分别用红色和蓝色表示。符号-表示已删除的核苷酸。
由于自交F1杂交稻中将减数分裂转化为有丝分裂和消除父本基因组都是可能的,我们通过同时编辑CY84中的OSD1、PAIR1、REC8和MTL4个基因来检验杂交稻诱导杂合固定而不额外杂交的可能性(图
剩余内容已隐藏,支付完成后下载完整资料

英语原文共 7 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[597712],资料为PDF文档或Word文档,PDF文档可免费转换为Word
课题毕业论文、外文翻译、任务书、文献综述、开题报告、程序设计、图纸设计等资料可联系客服协助查找。
您可能感兴趣的文章
- 莲NnWRKY基因在激素条件下的表达谱研究开题报告
- 使用转录组学分析阐明5-氨基乙酰丙酸在谷氨酸棒杆菌上的积累产生的生理效应外文翻译资料
- 基于网络药理学的水仙抗肺癌作用研究外文翻译资料
- 灵敏简便的仿生纳米酶免疫吸附比色法和表面增强拉曼效应检测 triazphos外文翻译资料
- LONELYGUY基因家族:从苔藓到小麦中生成植物活性细胞分裂素的关键基因外文翻译资料
- 葛根提取物可保护由过氧化氢引起的人视网膜色素上皮细胞死亡和膜通透性改变外文翻译资料
- 从人参中分离的内生细菌可促进生长、降低发病率并刺激人参皂苷生物合成外文翻译资料
- 拟南芥根表皮细胞命运的调控:多重反馈环的重要性外文翻译资料
- 根毛模式及形态发生的调控及可塑性外文翻译资料
- 拟南芥根表皮细胞分化的基因调控网络外文翻译资料

