裸子植物花粉萌发与花粉管发育
作者:Maria Breygina * , Ekaterina Klimenko and Olga Schekaleva
单位:Department of Plant Physiology, Biological Faculty, Lomonosov Moscow State University, 119991 Moscow, Russia; kleo80@yandex.ru (E.K.); anny.shirly.ganbatte@gmail.com (O.S.)
* Correspondence: breygina@mail.bio.msu.ru; Tel.: 7-499-939-1209
摘要:
花粉萌发和花粉管生长是种子植物普遍存在的过程,但这些过程最早出现在裸子植物中,至今仍对其有性繁殖起着重要作用。然而,关于生殖生理学的主要数据是在开花植物上获得的,因此人们应该小心地将发现的模式推断为裸子植物。近年来。针叶树花粉的生理研究日益增多,已初步确定了针叶树花粉的特征及其与开花植物的相似性。本综述的主要部分是针叶树花粉的生理学研究。本文展开介绍了裸子植物花粉粒的主要性质、多样性和传粉策略。
关键词:植物繁殖;花粉萌发;花粉管生长;裸子植物;松柏;云杉;松属;雄配子体;极地增长
- 介绍
裸子植物是植物界的优秀代表。一方面,它们是具有原始解剖结构特征的古老植物;另一方面,它们完美地适应了自己的栖息地,由于它们惊人的体型和寿命,让它们成为许多生态系统中的优势物种,尤其是它们的生殖系统。它具有渐进式的特征,因为在这个类群中,首先出现的雄性配子体花粉粒减少,以及形成种子的能力。此外,这一群体仍然代表着各种各样的生殖模式、策略和关系。例如,配子体减少的程度各不相同,既有动物配子体也有虹吸配子体,繁殖过程都可以相对较快,持续数年。然而,植物学和进化方面的研究已经在其他文献中进行了足够详细的描述[1-4],而本文主要关注针叶树花粉萌发过程中积累的生理数据;在2005年[5]发表了对这个主题的翔实的概述。在这里,我们主要考虑过去16年积累的新数据,以及一些重要的方面,而这些方面在之前的回顾中没有关注
- 裸子植物花粉粒特征
在裸子植物的进化过程中,从动物配子到管状配子的转变,即通过不运动的配子而不是运动的精子进行受精,现在这两种选择都在这个分类群[1]中被发现。现存裸子植物有12个主要科83属。术语“针叶树”指的是组成松植物门的一组裸子植物。迄今为止,裸子植物的生理学研究几乎都是在针叶树(主要是松树和云杉,很少有柏树和冷杉等)上进行的,所有这些植物都具有典型的虹吸配子受精(siphonogamy)特征,即配子静止不动。针叶树花粉的生理学在下面几节中讨论。
裸子植物的雄配子体一般比开花植物的更复杂;它的发育和发芽较慢;它是通过三到五次有丝分裂形成的,在不同的类群中发生在不同的阶段——授粉之前或之后。因此,小孢子囊释放出的花粉由不同数量的细胞组成(图1):松树和银杏有4个细胞(2个原叶细胞、2个雄蕊细胞和1个管细胞),红杉有2个,杜松有1个[6-8]。所有情况下的雄性配子都是如此授粉后形成的。
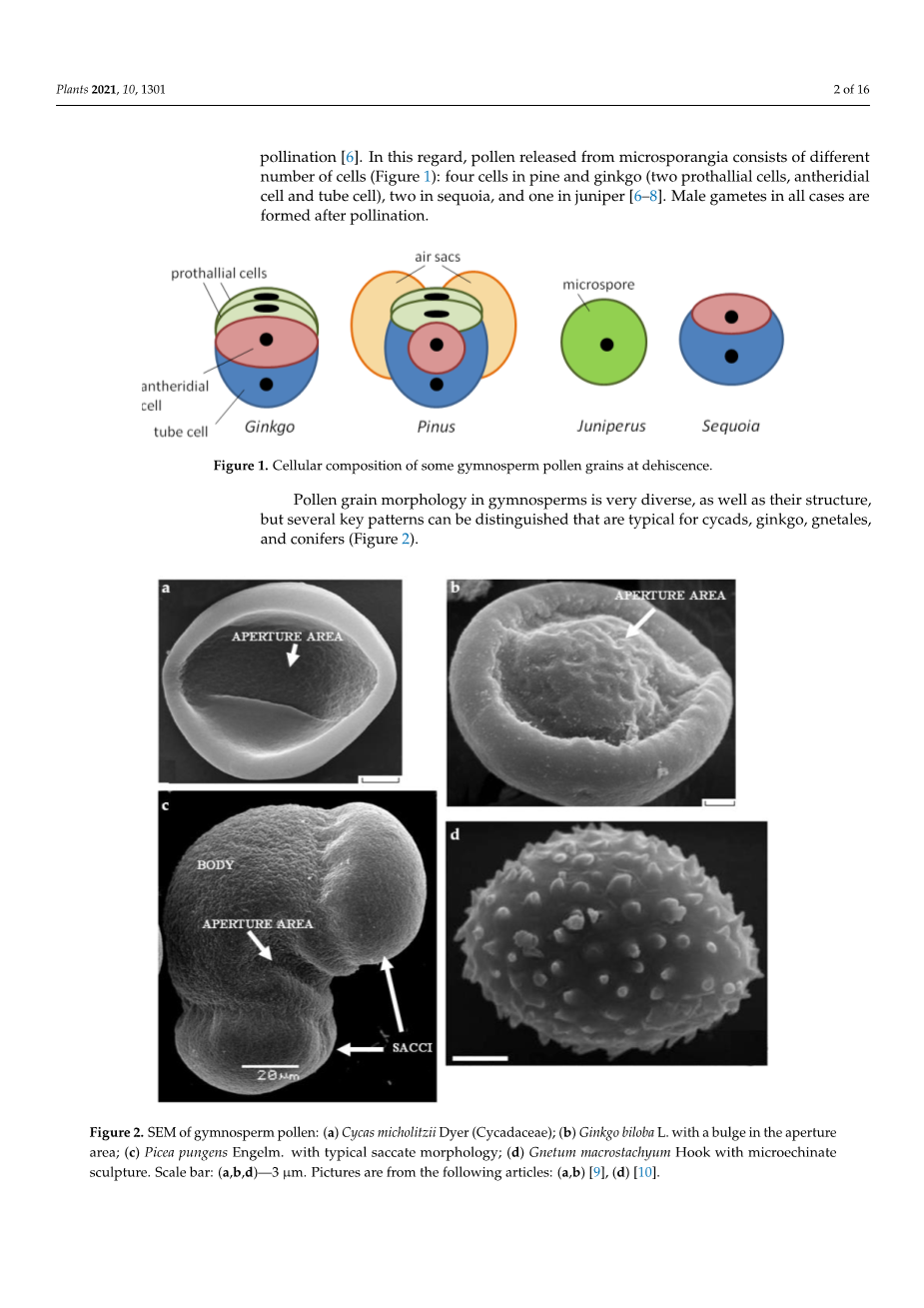
裸子植物花粉粒的形态和结构非常多样化,但可以区分出几个kev模式,这些模式在苏铁、银杏、叶虫和针叶树中是典型的(图2)。
配子体细胞被薄壁隔开,从外看,像开花植物一样,由内纤维素壁(内质)和大量的外孢粉壁(外质)保护。孔是特殊的区域,其中外壁较薄,内质通常增厚,沟道,或形成多层[12]。在许多情况下,花粉壁具有极性结构,裸子植物(如被子植物)小孢子的极性是在减数分裂过程中确定的。近端极(离四分体中心更近的那个)后来变得更凸,远端极变得更平,如银杏叶(图2b);外壁近侧较厚,大孔位于远侧。细胞学极性表现在细胞排列上:原叶细胞位于近端,大管细胞位于远端[13-15]。苏铁花粉粒表面的气孔几乎占到花粉粒表面的一半,外壁近端最厚,远端最薄,内壁较薄,外壁光滑(图2a)[16]。片麻岩中孔数和极性的问题还没有完全弄清楚。在磁囊草中,花粉是无孔的,内部极性,外壁是浮雕的(图2d)[10,17,18]。
在大多数松科植物中,花粉是囊状的,这意味着,它有囊(气囊)-由外壁形成的大的中空突起(图1和2c)。花粉囊位于花粉粒的远端,近端外壁增厚。气囊的存在与许多针叶树的授粉机制有关:花粉落在授粉滴上。这是接受胚珠分泌的液体,它形成一个大滴,上升到珠孔表面。花粉滴将花粉吸进胚珠中,在胚珠中萌发[19,20]。模型实验表明,花粉粒在传粉液滴中漂浮(由于花粉囊的存在)是花粉粒缩回至珠孔的有效机制;因此,气囊的作用不仅仅是通过风传递花粉。在配子体发育过程中,花粉囊的数量、花粉粒的大小和有丝分裂的数量都是变化的。裸子植物的花粉与被子植物不同,没有粘腻的富含脂质的花粉囊。
- 裸子植物授粉与花粉萌发
大约98%的裸子植物是风媒传粉的,这在很大程度上决定了它们的结构特征(例如,花粉粒和气囊的特定形状)和生理机能。据了解,几乎所有现代裸子植物的花粉,除了南洋杉科(Araucariaceae)和麻草属(Gnetum)的一些物种外,都具有高度脱水的特点,花粉可以覆盖很远的距离。与昆虫(主要是甲虫)的传粉在一些苏铁目和叶虫目物种中已经被描述过[17,24],包括化石,这可能是因为在古代苏铁的球果[25]中发现了粪化石。现存花粉粒的壁结构有显著差异。它们是由风携带还是由昆虫携带,即使是在同一个属内,麻黄就是这样证明的[171]在离体培养时,许多针叶植物的花粉在1-2天内萌发。许多物种已经制定了协议[26-28],允许有效的花粉萌发和长期监测其行为。针叶树花粉管的体外研究表明,针叶树花粉管与被子植物花粉管在细胞骨架组织、细胞器运动调控和胞内/胞外作用[5]等方面存在差异,我们将在下一节讨论。
在裸子植物中,花粉管出现在籽粒表面的位置并不像大多数开花植物那样是预先决定的。后者在大多数情况下有几个孔中的一个,旨在快速管出口[29]。根据拟南芥的数据,最佳的孔径数为3个[30]。在囊状针叶树花粉粒中,花粉管出现在囊之间,在与原叶状细胞相反的极点。本例的孔径区是外壁较薄的沟,其远端极有可能出现外壁破裂(图3a,c)[31]。然而,由于有两个这样的区域,一些花粉粒产生的不是一个而是两个管(图3a),这可以被认为是降低发芽率的一个特征,或作为一个潜力适应。在对蓝云杉花粉离体萌发的研究中,这种现象被描述为“双极性萌发”,它只发生在最适萌发培养基的情况下,显然,这不是一种获取额外营养的适应。如果花粉粒中没有气囊,但被极化,则管出的地方是与原叶细胞(麻黄或Welvitschia花粉)相反的一个相当宽的区域(图3b,d)[32]。单细胞花粉(杜松)中未发现极性的迹象,认为试管外观的位置尚未确定[6,33,34]。
在大多数裸子植物中,花粉粒到达雌球果后落在授粉滴上。液滴体积范围为10 nL至1 uL [36];大滴的水珠用肉眼是看得见的。没有授粉减少的物种的名单减少了。随着传粉研究的深入,包括通过系统发育定位[4]。一个小的或迅速消失的水滴经常被发现。授粉滴首先是一种外质液体。它含有无机物、碳水化合物和蛋白质,包括酶[20,33,36,37]。将传粉滴与已研究较多的被子植物的质外液(柱头和胚珠渗出液,花蜜)进行比较,发现它们的组成具有显著的相似性。在此基础上,认为花粉滴的作用不限于将花粉输送到胚珠。液滴酶,特别是几丁质酶,可能参与保护胚珠免受病原体侵害。本文还讨论了花粉萌发过程中液滴组分可能的调控功能。然而,所有这些假设都没有得到实验验证。
针叶树有不同的授粉策略[2,38]。松木和云杉的囊状花粉漂浮在传粉滴的表面,随着花粉被转移到珠心,在那里它发芽。在冷杉中,一种类似的授粉滴是由雨水或露珠收集的水分形成的。花粉通常在微柱管中萌发,花粉管向珠心生长。落叶松没有授粉滴,但微柱管充满卵珠分泌物。在这种分泌物的包围下,花粉水合、膨胀并脱落外壁。在这种形态下,花粉管漂到珠心,在珠心形成花粉管。这通常发生在授粉后的几个星期。在一些铁杉属植物和南红杉科植物中,没有授粉花粉滴落在靠近胚珠的圆锥形表面,并可能在那里停留数周。然后它发芽,形成一个长管,穿过珠孔进入胚珠,到达珠心。雌球果中花粉行为的多样性在文献中得到了广泛的讨论,因为它可以为授粉的进化提供重要的信息[2,20,22]。比较被子植物和裸子植物,我们发现后者的雄配子体授粉后往往需要覆盖较短的距离,但它的时间要长得多。雄配子体的萌发和生长在各个阶段都是缓慢的:针叶树花粉的水化通常发生在授粉后的第一天,花粉管在几天内出现,而开花植物的这一过程需要几分钟或几小时[5,39]。因此,云杉花粉管的生长速度约为20 um/h,与被子植物300 ~ 1500 um/h的生长速度形成鲜明对比。另外,针叶树的特征是花粉管不生长时有一段很长的休眠期。特别是,试管生长可以停止所需的时间,以完成雌性配子体发育长达一年。在这种情况下,花粉管在受精前几天恢复生长,并将精子输送到胚珠。例如,在松树和一些南洋槐花科植物中,花粉在授粉后不久就开始萌发,花筒进入珠心。冬眠期从仲夏持续到第二年春天。针叶松试管的休眠期始于胚珠的减数分裂,并在受精前几天结束。因此,对于不同的植物类群,从授粉到受精的时间从几周(柏科和松科的大多数物种)到一年(松科和一些南洋槐花科)不等。
- 针叶树花粉管的结构网络
4.1. 细胞骨架与细胞器运动
没有胼胝质塞在裸子植物花粉管,因此整个管充满一个数组的细胞质和细胞学上细分为顶点和身体(图4)。轴向微管和微丝束,管细胞的生殖细胞细胞核,无数造粉体,液泡,而其他的细胞器则局限在体内。透明区不含淀粉体,但富含线粒体和内膜成分,从顶端延伸20- 30um。这个区域(顶端)很容易与管体的其他部分(体)区分开来。花粉管顶端的小泡堆积呈新月形。樟子松(Pinus sylvestris)和云杉(Picea abies)花粉管的细胞器运动被描述为一个直接的喷泉,而不是反向的。被子植物花粉管中细胞器运动的不同方向,而裸子植物则与肌动蛋白微丝极性相反有关[40,43]。目前还没有裸子植物管中微丝取向的数据,但数学模型证实了这一说法[42]。
对云杉(Picea)花粉管生长调控进行了详细研究[44-48]。微丝的破坏使管状细胞完全停止生长。同时,顶端的囊泡运动无组织,从而破坏了顶端细胞壁的构建[46,47]。然而,微管的破坏也会阻碍花粉管的生长,破坏花粉管顶端的细胞器运动和肌动蛋白组织[40,48,49]。这一效应揭示了针叶树和被子植物花粉管的重要区别,其中微管运输雄性生殖单位,而其他细胞质结构的长距离运输主要依靠肌动蛋白细胞骨架[443,50-52]。此外,这种实验效应还改变了细胞器运动的方向:云杉[40]出现了被子植物特有的反向喷泉模式。值得注意的是,在花粉管[53]细胞内[Ca2 ]的调节实验中也观察到了类似的效果。由此可见,针叶树花粉管中的细胞器运动是由微管和肌动蛋白微丝[48]共同提供的。由Ca 离子控制(图5)。通过云杉花粉管[49]的线粒体运动可以说明肌动蛋白和微管蛋白细胞骨架之间的相互作用和功能连接。肌动蛋白和肌球蛋白直接移动线粒体,微管动力学影响肌动蛋白的组织,从而控制线粒体的速度、轨迹和位置。肌动蛋白骨骼的破坏也影响了栽培的云杉(Picea meyeri)花粉的蛋白质组学——在总共600个蛋白质中鉴定出80多个差异积累的蛋白质,并从功能上分为以下几类:信号转译、细胞骨架相关、细胞生长和碳水化合物代谢。Latrunculin处理极大地改变了高尔基体、线粒体和淀粉体的形态。细胞器结构的干扰与参与其功能的蛋白质的差异表达相结合。本研究揭示了花粉管中肌动蛋白细胞骨架破坏的复杂后果,包括蛋白质谱和细胞结构的整体变化[54]。
在花粉管生长过程中,离子转运体的极性分布、活性氧和细胞器得以维持。线粒体和其他细胞器沿肌动蛋白束(橙色)呈喷泉状移动。细胞质钙梯度在顶端最高(红色染色),pH梯度(蓝色染色,碱性染色),ROS顶端梯度(H2O2分子以星号表示),膜电位梯度(膜染色,红色标记去极化的根尖室)是生长管的特征。线粒体产生超氧自由基。钙信号转导:激酶样钙调素结合蛋白(KCBP)结合钙调素(CaM),并以Ca2 依赖的方式调节细胞内运动。
4.2. 细胞壁
针叶植物花粉管的细胞壁沉积模式与开花植物明显不同,但也有相似之处[5,45]。因此,在松木中,花粉管基部没有胼胝质,相反,花粉管顶端和较年轻的部分有胼胝质,在接近籽粒的地方消失。随后,利用细胞化学染色和单克隆抗体研究了14种裸子植物花粉管壁的大致组成。尽管有微小的差异,罗汉松科的花粉管。松科(Pinaceae)、杉科(Taxodia
剩余内容已隐藏,支付完成后下载完整资料

英语原文共 16 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[598078],资料为PDF文档或Word文档,PDF文档可免费转换为Word
课题毕业论文、外文翻译、任务书、文献综述、开题报告、程序设计、图纸设计等资料可联系客服协助查找。
您可能感兴趣的文章
- 莲NnWRKY基因在激素条件下的表达谱研究开题报告
- 使用转录组学分析阐明5-氨基乙酰丙酸在谷氨酸棒杆菌上的积累产生的生理效应外文翻译资料
- 基于网络药理学的水仙抗肺癌作用研究外文翻译资料
- 灵敏简便的仿生纳米酶免疫吸附比色法和表面增强拉曼效应检测 triazphos外文翻译资料
- LONELYGUY基因家族:从苔藓到小麦中生成植物活性细胞分裂素的关键基因外文翻译资料
- 葛根提取物可保护由过氧化氢引起的人视网膜色素上皮细胞死亡和膜通透性改变外文翻译资料
- 从人参中分离的内生细菌可促进生长、降低发病率并刺激人参皂苷生物合成外文翻译资料
- 拟南芥根表皮细胞命运的调控:多重反馈环的重要性外文翻译资料
- 根毛模式及形态发生的调控及可塑性外文翻译资料
- 拟南芥根表皮细胞分化的基因调控网络外文翻译资料

